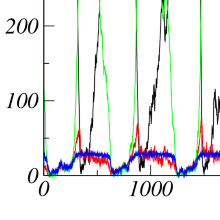
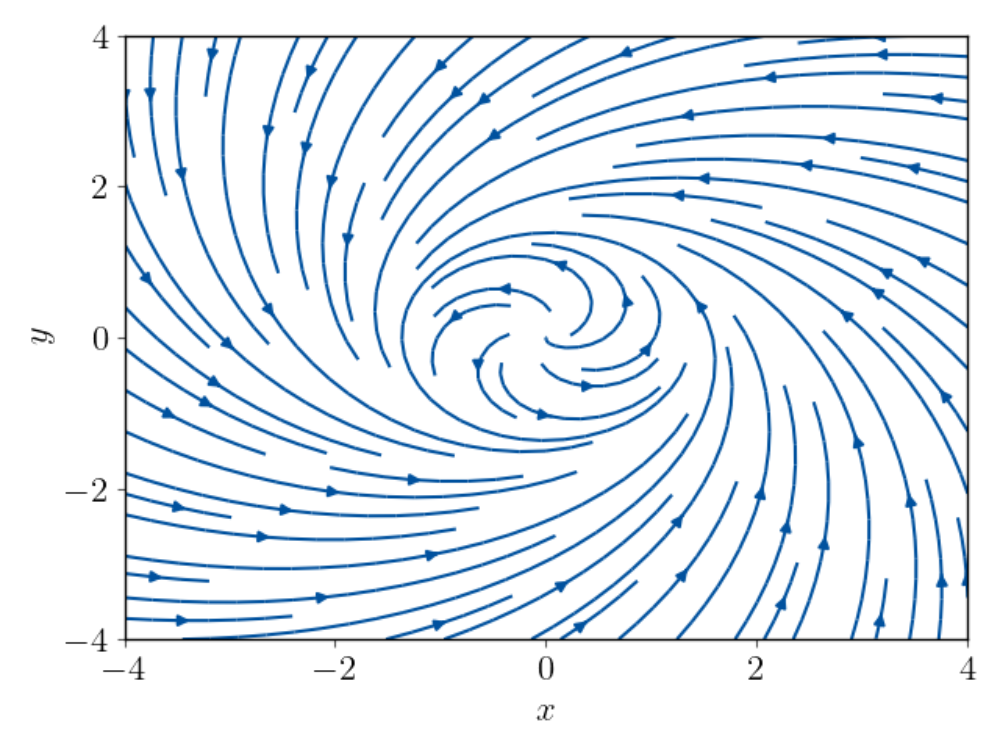
Thermodynamic cost of coherent oscillations
Biochemical oscillations are ubiquitous in living organisms. Sustaining these oscillations indefinitely in the presence of thermal fluctuations requires an external periodic signal. In the absence of such a stimulus, thermal noise effects allow only a finite number of coherent oscillations. This number is related to the free energy that is dissipated in the process, since strong affinity can suppress the desynchronizing effect of the fluctuations. In our work, we ask for the minimum amount of free energy consumption needed for a certain number of coherent oscillations.
Related publications:
- Universal minimal cost of coherent biochemical oscillations, L. Oberreiter, U. Seifert, and A. C. Barato, Phys. Rev. E 106, 014106, 2022, Abstract, Download
- Coherence of oscillations in the weak-noise limit, Benedikt Remlein, Volker Weissmann, and Udo Seifert, Phys. Rev. E 105, 064101, 2022, Abstract, Download
- Stochastic Discrete Time Crystals: Entropy Production and Subharmonic Synchronization, L. Oberreiter, U. Seifert, and A. C. Barato, Phys. Rev. Lett. 126, 020603, 2021, Abstract, Download
- Subharmonic oscillations in stochastic systems under periodic driving, L. Oberreiter, U. Seifert, and A. C. Barato, Phys. Rev. E 100, 012135, 2019, Abstract, Download
Phase transition in biochemical oscillators
We show that biochemical oscillations emerge through a generic nonequilibrium phase transition, where the control parameter is the thermodynamic force, which must be above a certain threshold for the onset of biochemical oscillations. They can occur in systems with a small number of molecules that display large fluctuations, thus precision is a relevant issue for biochemical oscillators. The number of coherent oscillations, which is the number of periods for which different stochastic realizations remain coherent with each other, is a standard measure of the precision of biochemical oscillators. The Fano factor associated to the thermodynamic flux has been proposed as an observable to quantify precision as it has a universal lower bound that depends solely on the thermodynamic force, which follows from the thermodynamic uncertainty relation. In the light of this phase transition, we show that the number of coherent oscillations quantifies precision reliably, while the Fano factor does not.
Related publications
- Phase transition in thermodynamically consistent biochemical oscillators, B. Nguyen, U. Seifert and A.C. Barato, J. Chem. Phys. 149, 045101 (2018), Abstract, Download
- Coherence of biochemical oscillations is bounded by driving force and network topology, A.C. Barato and U. Seifert Phys. Rev. E 95, 062409 (2017), Abstract, Download
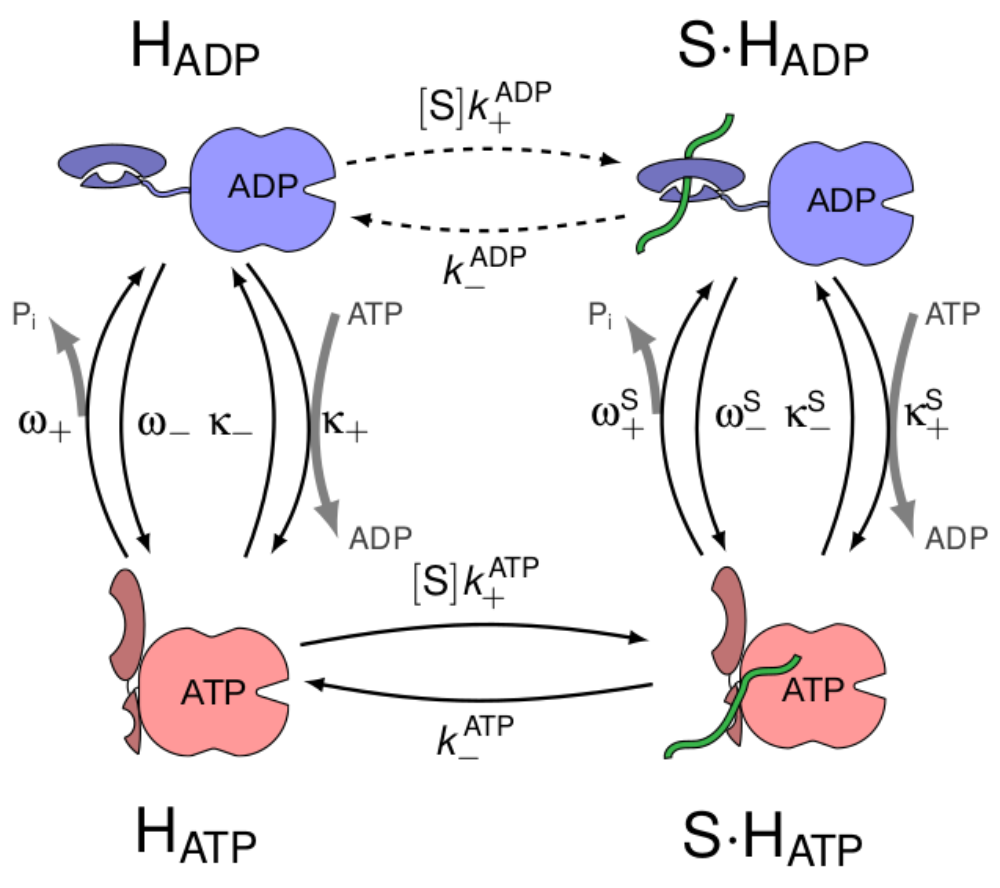
Thermodynamic bounds on the Hsp70
The 70 kDa heat shock protein Hsp70 has several essential functions in living systems, such as protecting cells against protein aggregation, assisting protein folding, remodeling protein complexes, and driving translocation into organelles. These functions require high affinity for nonspecific amino acid sequences that are ubiquitous in proteins. It has been recently shown that this high affinity, called ultra-affinity, depends on a process driven out of equilibrium by ATP hydrolysis. We have computed the thermodynamic bounds for ultra-affinity, and further shown that the same reaction scheme can in principle be used both to strengthen and to weaken affinities (leading in this case to infra-affinity). We show that cofactors are essential to achieve affinity beyond the equilibrium range. Finally, we propose that small GTPases can benefit from infra-affinity to optimize intracellular signal transduction.
Diffusion coefficient of Brownian ratchets
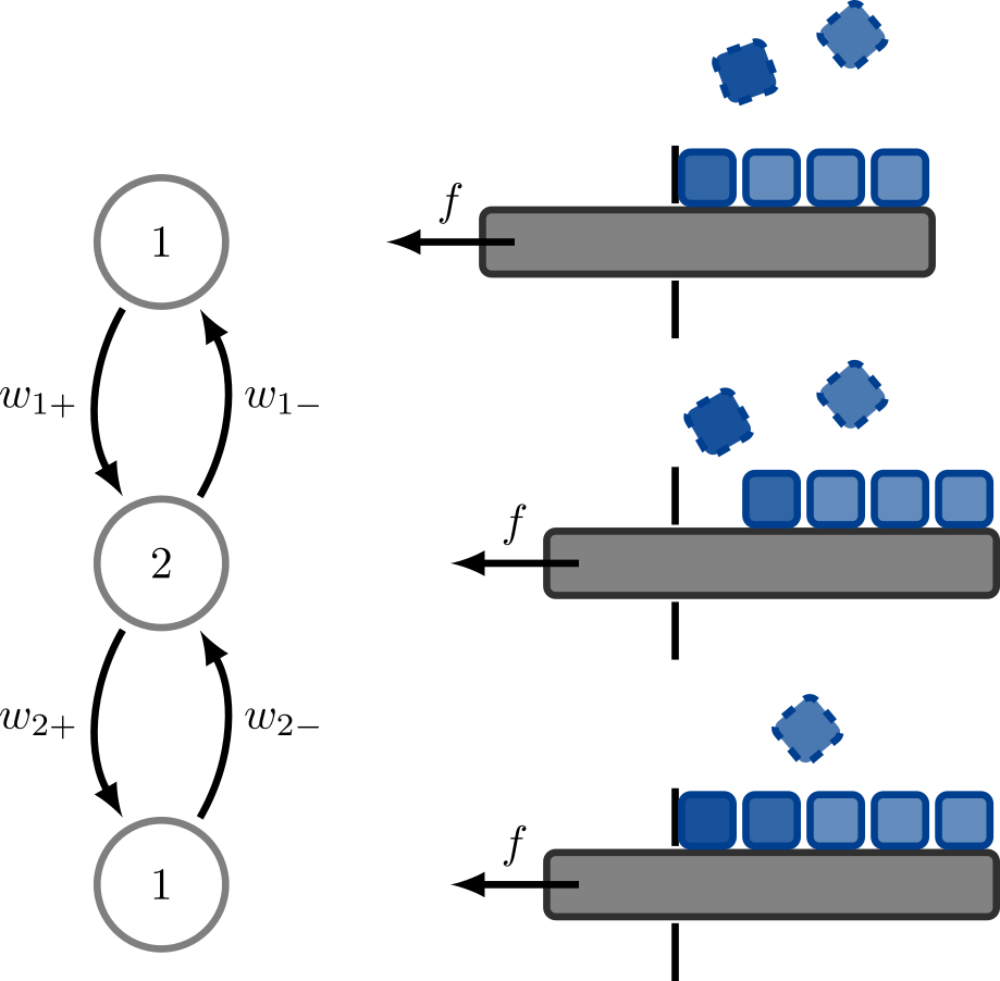
Initially introduced as a Gedanken experiment to understand how Brownian fluctuations can be used to extract work Brownian ratchets soon were used as model systems. For instance Brownian ratchets can be used to describe translocation of biopolymers like DNA through nanopores in cells in the presence of chaperones on the trans side of the pore. Chaperones can bind to the polymer and prevent it from sliding back through the pore. We study the diffusion coefficient and precision of such Brownian ratchets.
Sensory biochemical networks
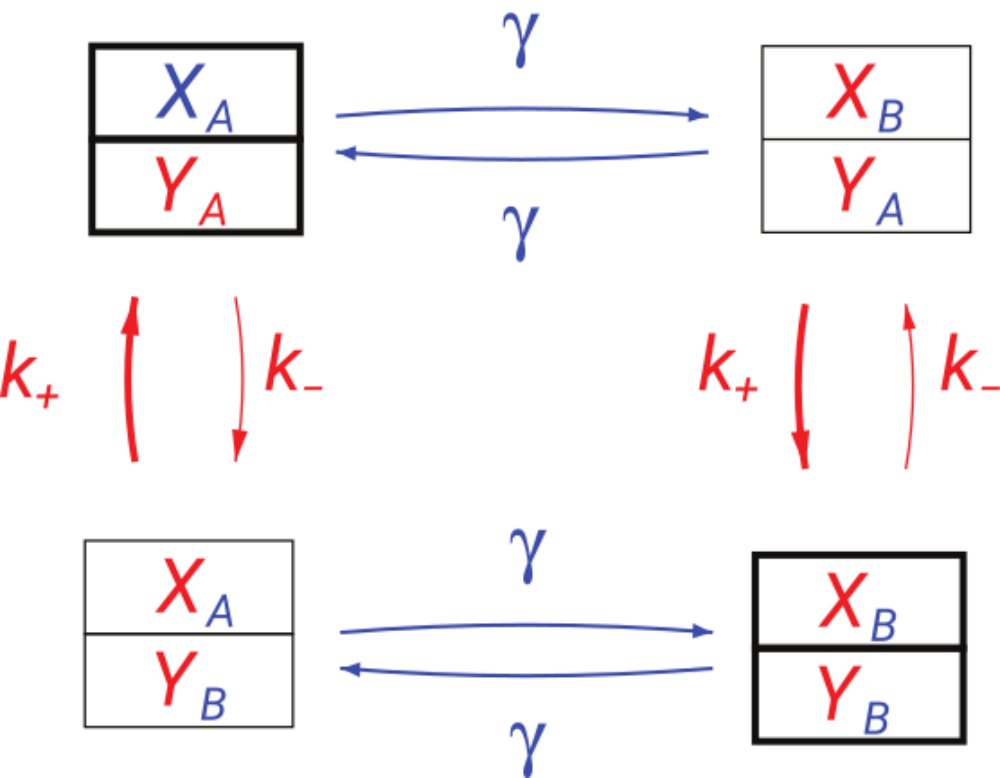
Acquiring and processing information about the instantaneous state of the environment is a prerequisite for survival for any living system. Sensory and signal transducting networks have evolved to achieve this task under a variety of external conditions. Maintaining any biochemical network, however, has a metabolic cost associated with its inherent non-equilibrium nature. The rate of mutual information characterizes the rate with which they acquire information about the changing external conditions. Comparing this rate with the thermodynamic entropy production that quantifies the cost of maintaining the network, we find that there is no universal bound restricting the rate of obtaining information to be less than this thermodynamic cost. On the technical level, in order to obtain the rate of mutual information we calculate Shannon entropy rates of non-Markovian time-series.